

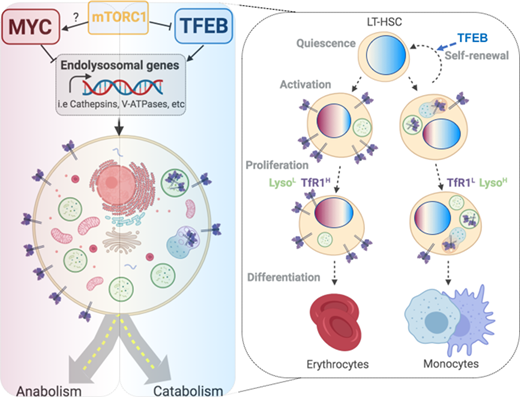
Human long-term hematopoietic stem cells (LT-HSC) residing at the top of the hematopoietic hierarchy must meet enormous daily demand (~10e11 cells daily) while also sustaining life-long maintenance of the stem cell pool through self-renewal. This hierarchical organization is widely thought to protect LT-HSC from exhaustion by maintaining them in a quiescent and undifferentiated state, activating only in response to microenvironment signals to generate highly proliferative but more short-lived populations including short-term HSC (ST-HSC) and committed progenitors. When called upon to exit this dormant state, HSC must respond and adapt their metabolism and nutrient uptake to meet increased bioenergetic demands for cell growth and differentiation. At the same time, the events underlying cellular and metabolic activation must also be suppressed to allow LT-HSC to re-enter quiescence and ultimately maintain the LT-HSC pool through self-renewal. Thus, proper sensing of cellular output demands must be coordinated with the cell cycle and metabolic machinery of LT-HSC to balance stem cell fates and maintain hematopoietic homeostasis. However, the regulatory circuits of this demand-adapted regulation of early hematopoiesis are largely unknown. The ability of cells to receive signals or take up nutrients depends on proteins that are embedded within the plasma membrane. These proteins move to the cell's interior through endocytosis and can be degraded in the lysosomes or rerouted back to the cell surface and reused. Moreover, lysosomes are the terminal catabolic stations of the autophagy pathway that is essential for preserving stem cell function through clearance of toxic cellular components. However, little is known about the regulation and role of lysosomes in the stem cell context. Here, we describe the unexpected finding that lysosomes, whose activity is intricately balanced by TFEB and MYC, are instrumental for regulating the stemness and differentiation properties of human LT-HSC. Furthermore, we found that TFEB, which is normally implicated in stress response, induces a constitutive lysosomal flux in unperturbed LT-HSC that actively maintains quiescence, preserves self-renewal and governs lineage commitment. These effects are accompanied by endolysosomal degradation of membrane receptors, such as the transferrin receptor 1 (TfR1), pointing to a role for TFEB in coordinating how LT-HSC sense environmental changes and initiate the earliest steps of their fate transitions and lineage commitment decisions. These transitions are regulated by a TFEB/MYC dichotomy where MYC is a driver of LT-HSC anabolism and activation and counteracts TFEB function by serving as a negative transcriptional regulator of lysosomes. Moreover, our findings further suggest that active suppression of TFEB and its downstream lysosomal degradation of TfR1 within LT-HSC is required for commitment along the erythroid lineage: activation of TFEB can abolish erythroid differentiation even after lineage commitment has occurred. In summary, we uncovered a MYC-TFEB-mediated dichotomous regulation of lysosomal activity that is required to balance anabolic and catabolic processes that ultimately impact human LT-HSC fate determination.
Takayanagi:Kirin Holdings Company, Ltd: Current Employment. Dick:Bristol-Myers Squibb/Celgene: Research Funding.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal